The extreme capsule in humans and rethinking of the language circuitry.
Makris, Nikos, and Deepak N. Pandya.
Brain Structure and Function213 (2009): 343-358.
1. 背景
伝統的に、ヒトの言語処理は大脳皮質の前頭-側頭頭頂領域を含むネットワークによって担われていると考えられてきた。特に、前頭葉のブローカ野、頭頂葉の角回、側頭葉のウェルニッケ野、そしてそれらを相互結合する弓状束 (AF: arcuate fascicle) は、言語の生成と表型に重要なネットワークの主要構成要素であると考えられてきた。これらの観察は臨床および病理学的研究に基づいている。近年、こうした考え方は画像研究によってさらなる支持を得てきており、特に拡散テンソル核磁気共鳴画像 (DT-MRI) の果たす役割は大きい。
マカクザルにおける研究によれば、ヒトの言語関連領域に相当する皮質領域は、他の結合路によって相互結合されているという。具体的には、前頭葉のBroca野に相当する領域は、側頭葉のWernicke野に相当する領域に、最外包 (EmC: extreme capsule) を介して結合されていることが示されている。また、実験研究によって、下部頭頂領域尾側 (角回) は、上側頭回 (STG: superior temporal gyrus) に相当する部位と、中縦束 (MdLF: middle longitudinal fascicle) という異なる連合線維によって結合していることが示されている。さらに、弓状束 (AF) は主に前頭葉の尾背側部 (area 8) と側頭葉のWernicke野に相当する部分の尾側 (上側頭回後部) を結合していることも示されている。これらの経路は、サルにおいて拡散スペクトル画像 (DSI: diffusion spectrum imaging) を用いることによっても示されている。
実験動物におけるこれらの経路のいくつかは、DT-MRIトラクトグラフィーとセグメンテーションを用いて、ヒトにおいても明らかにされている。すなわち、ヒトのMdLFは角回と上側頭回中部 (Wernicke野) を相互結合する一方で、AFは背外側前頭前皮質 (Areas 6, 8 and 46) と主にSTG最後部 (area Tpt) を相互結合することが示されている。古典的研究で示唆されていたように、Broca野とWernicke野の主要結合路はAFではないということは、動物における実験観察とヒトにおけるDT-MRIの両方が示している。では、Broca野とWernicke野を結ぶ解剖学的経路は、具体的になんなのであろうか。上で述べたように、サルにおける実験研究は、Broca野に相当する部位とWernicke野に相当する部位が、最外包によって相互結合されていることを示した。よって、人においてこの課題を解決するために、今回我々は5人の健常ヒト被験者に対してDT-MRIトラクトグラフィーを用いて最外包線維を可視化し、EmCがサルと同様の軌道および結合を持つのかどうかを評価した。
我々の結果は、最外包が長距離連合線維経路であり、鉤状束 (UF: uncinate fascicle)、最外包 (EC: external capsule)、MdLF、AF、上縦束 (SLF: superior longitudinal fascicle )II/III、下縦束 (ILF: inferior longitudinal fascicle) などの近接した線維束とは区別されるということを明確に示した。さらに、EmCは下前頭回 (Broca野) と上側頭回中-後部 (Wernicke野) の間に広がる線維であるということも示された。
2. 方法
我々はMRIを用いてEmC線維を定量的にトレースし、5人の健常右利きヒト被験者 (男性3人、女性2人、年齢23-24歳) においてその軌道を可視化した。DT-MRIに基づくトラクトグラフィーを用いた。皮質下構造 (前障や被殻など) のセグメンテーションと皮質区画化には、T1に基づく形態学的手法を用いた。
2-1. MRIプロトコル
Siemens Trio 3 T imaging system を用いたMRIを実施した。T1強調画像およびDT-MRI echo-planerプロトコル (60個の横断セクションから成る全脳横断拡散テンソルイメージング) を撮像した。
2-2. トラクトグラフィー
トラクトグラフィーは、線維経路の軌道の大部分を可視化できる。さらに、トラクトグラフィーは大脳皮質内の線維経路の相互関係を可視化でき、線維経路の区別を行うことを可能とする。トラクトグラフィーを用いて線維束を生成するためには、シードが必要である。一般に、シードは1つ以上のボクセルから成る。
2-3. シードの設定
シードは、ヒト脳の解剖学的知見や、実験動物の文献を用いて決定した。白質のT2-EPI (echo-planer image) の固有単位ベクトル (吻-尾側方向) をカラー符号化した冠状断画像に対して、EmCの "stem" portion に該当するボクセルを選択/ラベルした。選択されたボクセルがEmCのstem portionに該当するかを決定するために、2つの基準を用いた: (1) ボクセルのトポグラフィー (位置)、および (2) 組織の拡散特性の方向。冠状断では、EmCのstemは前障と島の間で、前後方向のテンソル情報を持つボクセルから構成される (図1)。前障と島の同定は、皮質下構造のセグメンテーションと皮質層の区画化を行った包括的研究の一部分として行われた。このステップによって、図1に示されるEmCのstemとECに該当するボクセルの選択に正確性と確度が増す。さらに、明確性、再現性、一般的利用可能性を期すために、個々のEmC (10半球) に対するシード点の定義を、Talairach座標システムで決定した。これは、各脳に対して、それぞれの冠状断スライスにおけるシードの重心をTalairach座標で決定することで行われた。

図1. (a) カラー符号化されたT2-EPI冠状断画像の固有単位ベルトルマップの代表例。(b) T2-EPIセクションに対応するT1強調画像。(c) 大脳半球の矢状断画像。(a) と (b) の冠状断画像は、(c) における黄色線のレベルで得られたものである。(b) の左半球には、皮質下構造と新皮質のセグメンテーションが示されている。(b1) は、関心皮質領域の区画化を行った上で、EmC、前障、ECの位置を示したものである。(a1) は、冠状断セクション (a) の拡大像である。EmC (緑ボクセル) が島 (茶ボクセル) と前障 (濃青ボクセル) の間のスペースの大部分を占めていることがわかる。(b) でも、EmCの位置が濃緑で示されている。また、 (c) では、EmCの軌道が吻尾方向に示されている。中央列の色付き球は、DT-MRIデータ解析で用いたカラー符号化スキームである。赤が右-左、緑が前-後、青が上-下方向である。(b) の黄色アスタリスクは前障の位置を示している。以下、略語の詳細。CGa cingulate gyrus, anterior; F1 superior frontal gyrus; F2 middle frontal gyrus; F3o inferior frontal gyrus, pars opercularis; FO frontal operculum; EC external capsule; EmC extreme capsule; INS insula; Put putamen; SC subcallosal cortex; TP temporal pole; L left; R right; A anterior; P posterior。

図2. 5人の被験者のEmCのシード点を、Talairach空間において示したもの。(a, b) 矢状断、(c) 軸位断、(d) 冠状断。前交連 (AC: anterior commissure) は0mm、後交連は-24mmに対応する。各被験者に対して、EmCシードが中央列のカラー符号に基づき色付けされている。
このトラクトグラフィーで観察された結果を確認するために、我々は近接する線維路とEmCを区別することを試みた。具体的には、EmCの周囲の線維路として、EC、UF、AF、SLF II/III、ILFを同定した。各線維路のシードの選択は、以下の通りに行った。ECのシードは前障の内側かつ被殻の外側 (図1b, b1) に、UFのシードは前頭側頭接合部と島限に設定した。AFとSLF I〜IILのシードはMakrisらと同様のものとし、ILFのシードは側脳室側下角の外側の側頭白質コアとした。EmCを近接するこれらの線維路と区別できれば、EmCが下前頭回と上側頭回を結ぶ長距離白質線維路であるということが、より高い確度で言えよう。
2-4. 最外包の同定
シードの選択に引き続き、トラクトグラフィーを実施した (図1c)。DTI Task Card ソフトウェア (MGH) を用いた解析を行った。このソフトウェアで用いられるアルゴリズムは、シード点から固有単位ベクトルの方向に追跡を行うというものである。
我々は、EmCのfractional anisotropy indexとして、異方性の平均値と標準偏差を計算した。さらに、EmCの体積を、ボクセル数とボクセルの体積の積として計算した。
各被験者に対して、DT-MRIに基づくトラクトグラフィーを用いて、EmCの軌道を決定した。トラクトグラフィーの結果に基づき三次元再構成を行った (図3)。さらに、DT-MRIと皮質区画化を組み合わせることで、EmCの皮質結合が推定された (図4)。また、EmCの周囲にある、EC、UF、AF、SLF II、SLF III、ILFの線維路の三次元再構成も行った (図5)。

図3. 10個の半球におけるDT-MRIに基づくトラクトグラフィーから得た、EmCの軌道を示す三次元描画像。
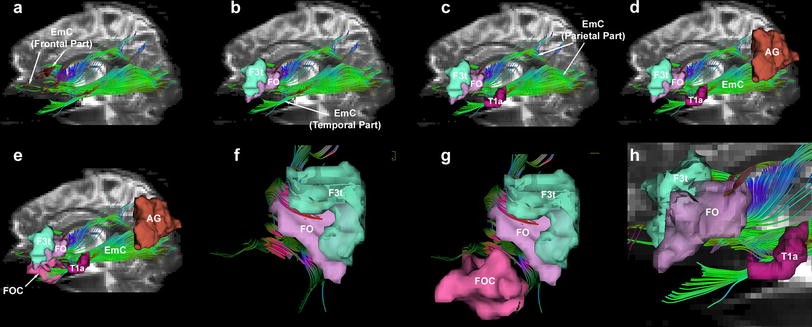
図4. EmCの軌道と、それが結合する皮質領域が、代表的被験者 (図3の1L) の描画シリーズに示されている (a-e)。EmCは前頭、側頭、頭頂部を持つと思われる (a-d)。前頭部の線維は、下前頭回皮質 (F3tとFO) と眼窩前頭皮質 (FOC) の中に存在する (b, e, f, g)。頭頂部の線維は下頭頂小葉 (AG: angular gyrus) の中に存在する (d, e)。側頭部の線維は上側頭回皮質 (T1a) の中に存在する (c, e, h)。

図5. (a) EmCを緑に、その他の線維束を他の色で同時に表示した図。DT-MRIトラクトグラフィーを用いることで、我々はMdLF、EC、UF、AF、SLF II、SLF III、ILFといった近接線維を、EmCと区別することができた。EmCはEC (ピンク) の外側に存在する。これら2つの線維路は、前障 (濃青, 赤アスタリスク) によって分離されている。UF (白) はEmCの腹側に、AF (黒)、SLF II (ターコイズ)、SLF III (青)はEmCの背側および外側部に位置している。ILF (赤) はEmCの腹側に存在する。(b) (a)において矢印で示したレベルにおける4つの冠状断セクションから成る。a-cは、同じ冠状断画像を異なる角度から見たものである。
3. 結果
我々は、fractional anisotropy (FA) と体積測定を行うことでEmCを同定し、トラクトグラフィーを用いることでその軌道を追跡した (図3)。次に、我々はシードをTalairach座標空間 (図2) にマップし、セグメンテーションと皮質区画化研究の結果を用いながら、皮質起源および終止部に至るまで、トラクトグラフィーの描画を行った (図4)。最後に、我々はEmCと近接する線維路であるEC、UF、AF、SLF II、SLF III、ILF をEmCと区別した (図5)。
3-1. トラクトグラフィーの観測結果
一般に、EmCの線維は下前頭回から上側頭回、さらには下頭頂小葉にかけて広がっているように見える。図4では、EmCの線維の軌道と、これらが結ぶ皮質領域が連続して示されている。EmCは、前頭部、側頭部、頭頂部の3つの部分から成るように見える (図4a-c)。図4a, bに示されるように、EmCの前頭部は下前頭回の中に存在する。図4fでは、EmCの線維がBroca野 (F3tとFO) に入っていくところが強調されている。なお、これらの皮質領域の位置と定義は、Center for Morphometric Analysis から引用したものである。図4e, gでは、EmCと眼窩前頭皮質の関係が強調されている。図4cでは、EmCの側頭部がSTGの前-中部 (赤で示されたT1a) に入っていくところが強調されている。図4hは、EmCと上側頭回のT1a野の関係性をさらに強調したものである。図4dは、EmCの頭頂部が下頭頂小葉 (角回またはAG) の中にあることを示している。図4eは、EmCの軌道全体を、Broca野 (F3tとFO)、Wernicke野 (T1aと上側頭回中部)、角回との関係とともに可視化したものである。また、トラクトグラフィーによって、近接する長距離線維路とEmCを区別することもできた (図5, 6, 7)。たとえば、図5では、SLF II (ターコイズ) とSLF III (青) は、EmC (緑) の背側の位置を占めている。同様に、EmCはAF (黒色) とも区別された。SLF II/IIIは主にEmCの上側を通り、外側前頭前皮質から頭頂葉に至るものであるが、AFの線維は背外側前頭前皮質から島の周囲をアーチ状に迂回して上・中側頭回の尾部に至るものである。加えて、EmCはMdLFとも区別された (図6, 黄)。

図6. 代表的症例 (図3の1L) のトラクトグラフィーの結果による、EmCとMdLFの解剖学的位置の比較。EmC線維は、MdLFの傍を下頭頂小葉に向かって尾側に走行する。これらの線維経路は上側頭回の尾側と下頭頂小葉白質の内部を隣り合って走行するが、bで示すように互いに区別可能である。尾側では、EmCの線維はMdLFの線維の内側に位置し、吻側では、EmCの線維は島 (INS) の内側壁に沿って走行して前頭葉に入る。EmCの線維のもう1つの群は、aで示すように、上側頭回の内部でMdLFの腹側を走行する。吻側では、MdLFの線維は上側頭回の内部を側頭極に向かって走行する。

図7. 代表的症例 (図3の1L) のトラクトグラフィーの結果による、EmC (緑)、EC (ピンク)、UF (白) の軌道の比較。aおよびa'で示すように、UFはEmCの腹側に位置する。さらに、bで示すように、EmCはECの外側を走行する。また、EmCは前障の外側に位置するのに対し、ECは前障の内側に位置する。
これらの2つの経路は、上側頭回白質の中を互いに並行して走行しているが、位置と広がりによって区別可能である。MdLFは側頭極と角回の間に広がっているのに対し、EmCはより内側に位置し、前頭前野の下部および腹側部と上側頭回から下頭頂小葉にかけて広がっている。EmCは、側頭極から後頭葉にかけて吻尾方向に広がるILFとも区別可能である。さらに、図7に示すように、EmCはUFとも区別可能である。UFは側頭極領域に存在し、腹側前頭前皮質に広がる (図7a, a')。これは、EmCの内側に位置する。最後に、図7bおよびb'に示すように、EmCはECとも区別可能であった。ECはEmCの内側に位置し、これらの2つの線維経路は前障を介して存在している。
3-2. EmCの定量的解析
我々は、5人の被験者に対して、fractional anisotropy (FA) 値の平均値と、FAの対称性係数およびEmCの体積を測定した。FAの左/右それぞれの平均値は、0.48/0.45であり、総合平均は0.46であった。FAの対称性平均は、より左方寄りであった (7.3%)。FAの値は、軸索密度や方向一貫性、髄鞘化といった、線維経路の生物学的特性を反映している可能性がある。FAの低値は脳卒中部位で報告されており、おそらく軸索変性やグリオーシスを反映している。一方FAの高値は脳卒中部位の周囲の白質で報告されており、おそらく白質再構成を反映していると考えられている。EmC体積の左/右それぞれの平均値は、15.93/16.27 cm3であり、対称性平均は、より右方寄りであった (4%)。EmCを生成するために用いたシードは、EmCのサイズの平均2.8%にすぎなかった。
4. 考察
Marchi法を用いたサルにおける過去の実験的研究において、EmCの軌道が明らかにされた。こうした研究によれば、EmCは連合線維経路であり、前頭葉、島、側頭葉、頭頂葉皮質から発する線維を含んでいる。前向性アイソトープトレーサを用いたマカクザルにおける実験では、EmCは中側頭回と腹側前頭皮質 (尾側眼窩前頭皮質を含む) を結ぶ長距離連合線維とされた。EmCの大部分は前障と島の間に一している。さらに、拡散スペクトラムイメージング技術を用いた研究において、サルのアイソトープ注入研究の結果の妥当性が示された。他の実験研究では、EmCが上側頭回中部および吻側部 (areas paAIt, TAa, TS3) と、隣接する上側頭溝皮質 (TPO)、側頭平面、島の吻側部、帯状回吻側部 (areas 24/32)、腹外側前頭前皮質 (ventral area 9/46) の尾側、前頭極皮質 (area 10)、そして眼窩前頭皮質尾側と area 45 を結合する線維を含んでいることが示された。EmCの大部分は、上側頭領域の中部 (ヒトのWernicke野に該当する部分)、前頭前野の area 45 (ヒトのBroca野に該当する部分) を結合しているように思われる。したがって、EmCはMesulam (1998) が言語情報伝達中心と呼んだ重要な経路に該当すると考えられる。
ヒトにおける最外包線維の経路と、その機能的役割は未だよくわかっていない。Dejerine (1895) は、最外包が外包からの線維のみならず連合線維を含んでいると報告した。Crosbyらによれば、最外包は「前頭葉と側頭葉、頭頂葉と側頭葉、そしてこれらの領域と島皮質を結ぶ多数の連合線維を含んでいる」主要な連合線維路である。
DT-MRIを用いた今回の研究では、我々はEmCが島と前障の間に位置する長距離連合線維であり、上側頭回中部と下前頭回および眼窩前頭回の間を走ることを示した (図3)。この線維路は、尾側は下頭頂小葉に向けて、他の線維経路 (MdLF) に沿いながら上側頭回白質の内部を走行する (図6)。このMdLFの線維は、下頭頂小葉 (BA 39) を上側頭領域吻側部および中部 (Wernicke野) と結合するとわかった。EmCとMdLFは上側頭回の尾側と下頭頂小葉白質の内部を平行しながら走行するが、これら2つの線維経路は互いによく区別可能である (図6)。尾側では、EmCの線維はMdLFの線維の内側に位置する。島後部のレベルでは、EmC線維は島内側壁に沿って前頭葉に侵入する (図6)。一方で、MdLF線維は上側頭回の内部にとどまり、側頭極に向かって走行する (図6)。また、我々はその他の線維経路とEmCを区別することもできた。すなわち、EC、UF、AF、SLF II、SLF III、ILF である (図5)。特に、図7bおよびb'で示すように、EmCが前障の外側に位置するのに対し、ECは前障の内側位置を占める。UFはEmCの腹側に存在する (図7a、a') が、AF、SLF II、SLF IIIはEmCの背側かつ外側の位置に存在する。ILFはEmCの腹側位置に存在する (図5)。
EmCは上側頭回中部 (Wernicke野) と下前頭領域 (Broca野) を結ぶ線維を運ぶ長距離連合線維経路であるように思われる。このため、言語処理に関連している可能性がある。この提案は、言語回路に関連する経路についての伝統的な考え方に異論を唱えるもののように思われる。
4-1. 言語回路を再考する
Wernickeは、弓状束はBroca野とWernicke野を相互結合するという点で言語に関連する主要な線維経路であると提唱した。この推測は、言語の生成と多様な失語疾患の解釈にあたっての主要な考え方であった。GeschwindはWernickeの考え方を改めて強調し、弓状束の言語に関する役割を重要視した。さらにGeschwindは、言語疾患の解明にあたり、下頭頂小葉 (特に角回) の重要性を提案した。近年、CataniらはDT-MRIトラクトグラフィーを用いて前頭-頭頂-側頭線維経路、特に弓状束の軌道を詳細に検討した。彼らの考え方は、Wernicke-Geschwindのスキーマに概ね合致していた。彼らの観測に基づけば、弓状束の線維は主に3つの下位区分に分けられる。すなわち、前頭葉のBroca野に該当する領域と側頭葉のWernicke野に該当する領域を結ぶ長いセグメント、Broca野と頭頂葉のGeschwind野 (角回) を結ぶ前方セグメント、そして角回とWernicke野を結ぶ後方セグメントである。他のDT-MRI研究では、上縦束-弓状束線維システムの軌道がさらに詳細に明らかにされた。この研究によれば、上縦束-弓状束経路は4つの下位要素に分けられた。背側前頭-頭頂線維はSLF Iと名付けられ、中部前頭-頭頂線維はSLF II、そして腹側吻側線維はSLF III と呼ばれた。島の尾側部をアーチ状に回る独立した前頭側頭線維経路の存在も示され、これが弓状束 (AF) と名付けられた。ヒトにおけるこれらのDT-MRI観察結果とサルにおける実験研究の結果を比較すると、AFは前頭前野尾背側 (areas 8 and 46) と上側頭回尾側 (area Tpt) を結んでいることが示唆された。サルにおける実験観察、臨床観察、そしてヒトにおける構造的および機能的画像研究によって、上側頭回尾側 (area Tpt) は空間内の音の位置に関わり、より腹側の部分 (areas PaAIt, TS3 and TS2) は音の認知に関わることが示唆されている。これらの考え方や、ヒトにおける電気生理学的研究に基づくと、Wernicke野は主に上側頭回 (STG) の中部に位置すると考えられた。このため、ヒトにおいて弓状束がBroca野とWernicke野を結んでいるかどうかは未だ解決されていない疑問である。ヒトとは異なり、サルではこのような前頭側頭結合 (Broca野とWernicke野を結ぶ結合) は双方向性に最外包を通っていることが示されている。
ここから、EmCの言語に関する役割について疑問が浮上する。Cataniらは、角回 (Geschwind野) と上側頭回 (Wernicke野) の間は弓状束後部によって結合されると提唱した。彼らは、Geschwindによって提唱されたように、弓状束のこのセグメントが言語機能を持つと考えた。一方で、我々の近年の研究によれば、角回は弓状束ではなくMdLFによって上側頭回中部 (Wernicke野) と結ばれる。MdLFは、SLF II-弓状束システムの一部というよりも、独立した線維束と呼べるものである。上で述べたように、サルにおける実験研究は、弓状束が背尾側前頭前皮質と上側頭回尾側 (apea Tpt) を結んでいることを示した。これらの領域はどちらも、空間内の音の位置の同定に関わっているとされている。一方で、サルのarea 45を含む下部前頭前皮質は、上側頭回中部と上側頭溝皮質 (ヒトWernicke野に該当する部分) と、最外包によって結合されている。したがって、弓状束はおそらく主に空間内の音の位置の処理に関与しているのに対し、EmCとMdLFは言語に重要な役割を持っていると思われる。
ヒトとサルの両者において、SLF-AF線維システムは、EmCおよびMdLFと独立しているため、我々は言語に関連する構造と線維経路に関して代替的な仮説を提唱したい。これは、非ヒト霊長類における実験観察による推察に基づくものである。サルでは、area 45 (ヒトの三角部に該当) は上側頭回 (STG) 中部および上側頭溝 (sts) 皮質と、最外包を介して双方向性に結ばれている。これらの領域 (STGとsts) はそれぞれ、モダリティ特異的な聴覚的情報処理 (STG) とマルチモーダルな (sts) 情報処理に関与している。さらに、STGとsts皮質は、MdLFを介して頭頂葉のarea PG-Opt (ヒトでは角回に該当する領域) と結ばれていることが示されている。したがって、サルでは前頭-頭頂側頭回路は3つの皮質中心 (i.e., 前頭前野 area 45、上側頭回中部、下頭頂小葉尾側) によって構成されており、主に2つの長距離連合線維経路 (i.e., 最外包とMdLF) によって相互結合されていると言える。よって、ヒトにおける言語関連結合が次のように構成されていると考えるのは合理的である。下前頭回尾部 (Broca野) ⇔ 最外包 ⇔ STR (superior temporal region, i.e., STGとsts; Wernicke野) と、角回 ⇔ MdLF ⇔ STR。これらの相互結合は、言語表現 (EmCによって結ばれる側頭前頭結合) および言語理解 (MdLFによって結ばれる側頭頭頂結合) に関わっている可能性がある。この結合性は、図8a-dに示すように、本研究におけるトラクトグラフィーを用いた観察によって支持される。

図8. 3つの皮質中心 (i.e., 前頭前野 area 45、上側頭領域中部、下頭頂小葉尾部) から成る前頭-頭頂-側頭回路は、2つの主要な長距離連合線維経路 (EmCと MdLF) によって結ばれている。EmCによる結合は、パネルa〜dに示されている。下前頭回尾部と上側頭領域を結ぶEmC線維は、cとdに示すように、島の周りをループ状に走行する。このループの内側部 (EmCの前頭セグメント) は、下前頭領域尾部からSTRまで、島の内側壁に沿って走行し (d)、外側部 (EmCの側頭セグメント) は島の最尾側部で上側頭回に入り、下方かつ外側方向に島輪状溝下部に至るまでSTRの中を走行する (c)。EmC線維は上側頭白質を島限で抜けて、下前頭領域尾部と結ばれる (a, c)。MdLFは下頭頂小葉 (AG) から上側頭回白質の内部を経て側頭極に至る (a, b)。ここで示した前頭-頭頂-側頭回路は、伝統的なWernicke-Geschwindの言語スキーマの代わりまたは補完となり得るものである。下前頭回尾側 (F3t/FO: Broca野)、STR (T1a/T1p: Wernicke野)、AGおよびAF-SLF II/IILの位置関係がe〜hに示されている。AF線維は上側頭回の後端の尾側の周りでアーチを形成し、中側頭回 (T2aとT2p) に入る (f, g, h)。
上で述べた考え方はサルとヒトの両方における観察に基づいたものであるため、あくまで仮説に過ぎないということは指摘しておきたい。この提案の妥当性を評価するためには、機能的および臨床的な検討がさらに行われる必要がある。さらに、サルとヒトの機能的および構造的差異に基づくと、結合性の違いが存在する可能性も考えねばならない。したがって、ヒトではEmCとMdLFではなく、AFが主要な言語結合路である可能性も残る。また、EmCとMdLF、そしてAFのすべてが言語機能に寄与しているという可能性もある。
Rillingら (2008) による最近の比較研究において、AFが前頭皮質と中下側頭回を結んでいることが示された。彼らは、ヒトにおいて側頭領域とBroca野を結ぶ新しい結合路が成立し、語彙-意味処理および統語処理に関与している可能性を示唆した。Saurら (2008) によるfMRIを用いた研究では、EmCが言語機能において大きな役割を果たすことが指摘された。このデータでは、AFとSLF IIが言語における感覚-運動機能を果たす一方で、EmCはより高次の言語理解に関与していることが示唆された。これらの観察は、今回の我々の発見と結論を支持するものである。
4-2. 定量的解析
我々は、EmCのFAとFA対称性という2つの生物物理学的パラメータを測定した。FAは左で0.45から0.49、右で0.43から0.45であった。EmCのFA値は、ヒトやサルでは得られていないため、他の研究と比較することはできない。しかし、FA値は、正常ヒト脳の異なる線維路に関する他の研究で報告されたFA値と類似していた。PierpaoliとBasserは皮質下白質におけるFA値0.46を報告している。Klingbergらは、側頭頭頂部白質におけるスケールなしのFA値の平均が0.38から0.59であることを示した。また、帯状束 (CB)、上縦束 (SLF)、後頭前頭束 (OFF)、MdLFの先行研究では、CBのFA値は0.45から0.54、SLF (4つのサブコンポーネントをすべて合わせた) 全体のFAは0.44、OFFのFAは0.37、MdLFのFAは0.34であった。したがって、EmCのFAの結果は、ヒトの脳の他の線維経路のデータと概ね一致しているようである。EmCのFAは、7.33%の左方非対称性を示した。EmCの全体の体積は大脳の両側で約16cm3であった。このことは、EmCがSLF IIに匹敵するサイズの大きな連合皮質皮質繊維路であることを示唆している。また、EmC体積は、4%の右方への偏りを示した。
5. 結論
今回の研究では、我々はDT-MRIを用いてヒト生体における最外包の軌道を明らかにし、その結合を推定した。最外包は、下前頭回から上側頭回と下頭頂小葉にかけて広がる長距離連合線維経路である。我々は外包、MdLF、UF、AF、SLF II/III、ILFなどの近接する線維経路から最外包を区別した。この経路は下前頭回 (Broca野)、上側頭回中部 (Wernicke野)、下頭頂小葉 (角回) を結んで言語領域の中に存在することから、言語機能において突出した役割を持つ可能性がある。本研究は、最外包とMdLFを含む、言語回路の推定的代替経路を実証するものである。
6. 感想
久しぶりの投稿になりました。ここ数ヶ月忙しすぎたのですが、徐々に余裕が出てきたのでようやく記事書けました。最外包は島皮質下にあって前障の上にある白質のことですが、島皮質下を含む病変で臨床的に失語症状が見られるような気がしていて、いったいどことどこを結ぶ白質路なんだろうと思って読み始めたのがきっかけでした。
けっこうわかりにくい書き方だったので理解に苦しんだ部分も多かったのですが、著者のロジックは、「Broca野とWernicke野とGeschwind野は言語に大事」なので、「それらを結ぶ白質路を調べたら、言語に重要な白質路がわかるでしょう」ということなんだと思います。そして、「古典的にBroca野とWernicke野を結ぶとされていた弓状束は、実は背外側前頭全皮質と上~中側頭回尾部を結ぶもの」であり、一方で「最外包を通る白質はBroca野とWernicke野を結んでいました」と主張したいのだと思います。
彼らがトラクトグラフィーで明らかにしたのは、最外包を通る線維は、① 前頭部と側頭部と頭頂部に分けられ、② 前頭部はBroca野から島の内側壁に沿って後方に走行し、③ 側頭部は島の最尾側部でターンして上側頭回内部に入り、下方かつ外側方向に走行してWernicke野に至り、④ そこから上側頭回内を島輪状溝下部に至るまで走行し、上側頭白質を島限で抜けて、Broca野に戻る、ということです。つまり、最外包を通る線維はBroca野とWernicke野を2通りの走行経路で結んで、ループを形成しているということです。また、頭頂部についてはなぜかあまり触れられていなかったのですが、おそらく先ほど述べた2通りの経路の両方を通って、Broca野とGeschwind野が結ばれているということなのでしょう。この「2通り」というのがおそらく大切で、最外包は下前頭後頭束と鉤状束など2種類の線維経路が通る白質である可能性が高いと思われます。ここから予測されることって結構あると思うのですが、残念ながらこの論文にはそこまで書かれていませんでした。もう少しちゃんとdiscussion書いてほしかったなあ...。